
CECILIA I. CLOSS1, 2, MARTÍN A. RUIZ DIAZ1, ALBERTO M. CAFFERATA3, 4,
DAMASIA BECÚ-VILLALOBOS5, JUAN P. NOGUEIRA1, 2, 4
1Asociación Formoseña de Endocrinología y Metabolismo, 2Centro de Investigación en Endocrinología, Nutrición y Metabolismo (CIENM), Facultad de Ciencias de la Salud, Universidad Nacional de Formosa, 3Prevención Cardiovascular, Sanatorio Finochietto, 4Facultad de Medicina, Instituto Universitario de Ciencias de la Salud, Fundación H.A. Barceló, 5Instituto de Biología y Medicina Experimental (IBYME-CONICET-FIBYME),Buenos Aires, Argentina
Resumen En la diabetes mellitus tipo 2 el aumento en la producción de quilomicrón en el estado post-prandial
se asocia a mayor riesgo cardiovascular. La evidencia actual posiciona al enterocito como actor principal en la dislipemia de la diabetes mellitus tipo 2 debido a que aumenta la producción de apolipoproteína B-48 en respuesta a una elevación de ácidos grasos libres y glucosa. El metabolismo del quilomicrón se encuentra regulado por múltiples factores independientes además de la ingesta de grasa alimentaria. Entre estos factores se destacan las hormonas intestinales, como el péptido similar al glucagón tipo 1 que disminuye la producción de apolipoproteína B-48 y el péptido similar al glucagón tipo 2 que la aumenta. Por otro lado, la insulina inhibe de forma aguda la producción de quilomicrón en el sujeto sano mientras que en la diabetes mellitus tipo 2, este efecto está ausente. La comprensión de los factores reguladores emergentes de la secreción de quilomicrón permite vislumbrar nuevos mecanismos de control en su metabolismo y aportar estrategias terapéuticas focalizadas en la hiperlipidemia posprandial en la diabetes mellitus tipo 2 con la reducción del riesgo cardiovascular.
Palabras clave: diabetes mellitus tipo 2, quilomicrones, apolipoproteína B-48, dislipidemias
Abstract. Role of the enterocyte in type 2 diabetes mellitus associated dyslipidemia. In type 2 diabetes
mellitus there is an overproduction of chylomicron in the postprandial state that is associated with increased cardiovascular risk. Current evidence points out a leading role of enterocyte in dyslipidemia of type 2 diabetes mellitus, since it increases the production of apolipoprotein B-48 in response to a raise in plasma free fatty acids and glucose. The chylomicron metabolism is regulated by many factors apart from ingested fat, including hormonal and metabolic elements. More recently, studies about the role of gut hormones, have demonstrated that glucagon-like peptide-1 decreases the production of apolipoprotein B-48 and glucagon-like peptide-2 enhances it. Insulin acutely inhibits intestinal chylomicron production in healthy humans, whereas this acute inhibitory effect on apolipoprotein B-48 production is blunted in type 2 diabetes mellitus. Understanding these emerging regulators of intestinal chylomicron secretion may offer new mechanisms of control for its metabolism and provide novel therapeutic strategies focalized in type 2 diabetes mellitus postprandial hyperlipidemia with the reduction of cardiovascular disease risk.
Key words: type 2 diabetes mellitus, chylomicrons, apolipoprotein B-48, dyslipidemias
Recibido: 8-IX-2017 Aceptado: 13-III-2018
Dirección postal: Juan P. Nogueira, Av. Dr. Luis Gutnisky 3200, 3600 Formosa, Argentina
e-mail: nogueirajuanpatricio@gmail.com
La principal causa de muerte de los sujetos con diabetes mellitus tipo 2 (DM2) es la enfermedad cardiovascular, principalmente por cardiopatía isquémica. La base fisiopatológica de esta entidad es la aterosclerosis acelerada asociada a la dislipidemia que presentan los sujetos con esta condición. La dislipidemia asociada con la DM2 se caracteriza por hipertrigliceridemia en ayunas, descenso del colesterol asociado con lipoproteínas de alta densidad (HDL-C), presencia de colesterol asociado a lipoproteínas de baja densidad (LDL-C) pequeñas y densas e hiperlipemia posprandial. Se considera que cada uno de estos 4 parámetros constituye un marcador de riesgo cardiovascular independiente1, 2.
Se ha demostrado mediante estudios epidemiológicos y de intervención que el principal factor de riesgo lipídico es el LDL-C. Se observó en sujetos con DM2 tratados con estatinas que el riesgo cardiovascular no desaparece totalmente en presencia de niveles bajos de LDL-C. A esta situación se denomina riesgo cardiovascular residual y la hipertrigliceridemia posprandial representa un factor importante de esta condición3.
El intestino delgado es un órgano vital para la homeostasis de los triglicéridos posprandiales provenientes principalmente de la vía alimentaria. Se ha demostrado que el intestino es capaz de realizar lipogénesis de novo y mediante la interpretación de sus mecanismos se podrían identificar blancos terapéuticos de utilidad en la dislipidemia asociada con la DM2.
El objetivo de esta revisión es actualizar sobre el rol activo del enterocito en la dislipidemia de la DM2.
Absorción y metabolismo intestinal del colesterol
La proteína 1 similar a Niemann-Pick C1 (NPC1L1) es un transportador de esteroles que se localiza en la membrana apical del enterocito y puede facilitar la captación de colesterol al promover el pasaje de esteroles a lo largo de la membrana del borde en cepillo y por endocitosis vesicular4. A su vez, se demostró que dos miembros de la familia de receptores basureros, como el receptor scavenger clase B tipo I (SR-BI) y el cluster determinant 36 (CD36) median la absorción de colesterol libre en la membrana del borde en cepillo del enterocito.
Se comprobó la existencia de un heterodímero de la superfamilia de transportadores ATP binding cassette G5/G8 (ABCG5/G8) que promueve la excreción de colesterol y fitoesteroles desde el enterocito a la luz intestinal. Existe una correlación negativa entre la absorción del colesterol y la expresión de ABCG5/G85. Los efectos regulatorios combinados de NPC1L1 y el ABCG5/G8 juegan un papel crítico en la modulación de la cantidad de colesterol que alcanza los vasos linfáticos desde la luz intestinal.
Se estima que la vía biliar contribuye en un 17% a la eliminación de esteroles del cuerpo. Se ha demostrado que existe una vía de eliminación del colesterol independiente de la excreción biliar clásica. Se postula que el enterocito captaría el colesterol de las partículas de HDL y LDL circulantes a través de su membrana basolateral para luego excretarlo a la luz intestinal por el heterodímero ABCG5/8 o por la proteína ATP binding cassette B1a/b6. Este mecanismo de excreción trans-intestinal del colesterol (ETIC) podría contribuir aproximadamente en un 33% a la cantidad total de esteroles neutros excretados por el organismo (Fig. 1)7.
Absorción y metabolismo de los ácidos grasos libres y los triglicéridos
Como se muestra en la Fig. 1, el CD36 mediaría la absorción de ácidos grasos de forma aislada o en conjunto con la proteína fijadora de ácidos grasos de membrana, la cual acepta los ácidos grasos de cadena larga en la membrana celular para incrementar su concentración local. Esto podría ayudar al CD36 a transportar de manera activa a los ácidos grasos a través de la membrana apical del enterocito8.
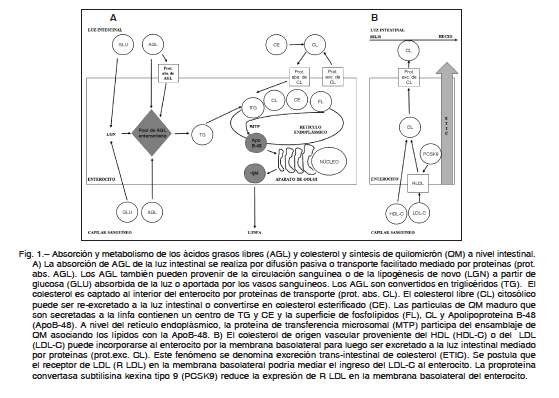
Una vez que se encuentran en la cara interna de la membrana del enterocito, los ácidos grasos se unen a la proteína fijadora de ácidos grasos citoplasmática antes de ingresar a las vías metabólicas. Estos ácidos grasos podrían ser utilizados para formar ésteres con la acil-CoA, reacción catalizada por la proteína transportadora de ácidos grasos 4 a nivel del retículo endoplasmático9.
Tanto los ésteres de acil-CoA con ácidos grasos como el monoacilglicerol se utilizan en el retículo endoplasmático liso para la síntesis de diacilglicerol y triglicéridos. La vía del monoacilglicerol contribuye en un 75%-80% en la síntesis de triglicéridos de origen intestinal. Por otro lado, la glucosa se transporta al interior del retículo endoplasmático liso y contribuye a la síntesis de triglicéridos por la vía del glicerol fosfato10.
Síntesis de quilomicrón
Los triglicéridos participan en la formación de QM que también requiere la presencia de Apolipoproteína (Apo) B-48 (ApoB-48) y de la actividad de la proteína de transferencia microsomal (MTP) a nivel del retículo endoplasmático. La MTP es un heterodímero compuesto por dos subunidades, una subunidad P de 58 kDa y una subunidad M de 97 kDa (Fig. 1). La MTP interactúa con los lípidos y los transfiere hacia la ApoB-48 naciente. Esto genera un cambio en la configuración estructural de la ApoB-48 haciéndola receptiva para aceptar más lípidos. Se demostró que la región promotora del gen de la MTP contiene un elemento de respuesta a la insulina mediante el cual dicha hormona inhibe la expresión de esta proteína11. A su vez, esta región promotora también contiene un elemento de respuesta a esteroles, y se observó que el descenso de colesterol intracelular reduce el contenido de ARNm de MTP12. En la maduración del QM participan tanto la apolipoproteína C-III13 como la apolipoproteína A-IV14; ambas apoproteínas estimulan el ensamblaje de triglicéridos a la partícula madura de QM. Se debe mencionar que si no se produce la lipidación de la ApoB-48 dicha apoproteína es degradada15. El número de ApoB-48 se correlaciona con el número de partículas de QM y de sus remanentes, los cuales son potencialmente aterogénicos16.
Una vez sintetizado el QM, se puede observar que su centro se conforma por triglicéridos y colesterol esterificado y la superficie de esta lipoproteína se encuentra formada por una monocapa de fosfolípidos (principalmente fosfatidilcolina), colesterol no esterificado y las apolipoproteínas entre las que se incluyen la ApoB-48, ApoA-I, ApoA-II, ApoA-IV y ApoA-V17.
Regulación de la síntesis de quilomicrón
En el esquema de la Fig. 2 se muestran los factores que modulan la producción de QM.
Ácidos grasos libres. El contenido de lípidos en la luz del intestino regula los niveles de CD36 el cual inicia una cascada de señalización que conduce a un incremento de proteínas clave en la síntesis de QM como la ApoB-48 y la MTP18, 19.
Se demostró que el aumento de ácidos grasos libres en el plasma mediante la administración de una emulsión de triglicéridos produjo un incremento en la tasa de producción de ApoB-4820. Esto demuestra que el ensamblaje de QM se produce también gracias al aporte de sustratos por vía sanguínea. El mecanismo adicional por el cual los ácidos grasos libres estimulan la producción de QM se debería a una mayor secreción de apolipoproteína C-III, mayor estabilidad de la ApoB-48 y su menor degradación intracelular21.
Colesterol. El ensamblaje de QM también se ve influenciado por la disponibilidad de colesterol presente en el enterocito. Se demostró en células CaCo-2 que el exceso de colesterol libre reduce la síntesis de novo de colesterol y estimula su esterificación. Esto resulta en un incremento en su almacenamiento en forma de gotas lipídicas citoplasmáticas y un aumento en la transferencia de ésteres de colesterol hacia la ApoB mediada por la MTP22. Como se mencionó previamente, la región promotora del gen de la MTP contiene un elemento de respuesta a esteroles que promueve la expresión de MTP ante un aumento de los niveles celulares de esteroles y suprime su expresión cuando estos se reducen.
Proproteína convertasa subtilisina kexina tipo 9 (PCSK9). La PCSK9 es una proteína encargada de reducir la expresión del receptor de LDL. Además, esta proteína cumple otras funciones. En un experimento realizado con células CaCo-2 la administración de PCSK9 exógena se asoció a una disminución de la expresión del receptor de LDL en la membrana baso-lateral, en conjunto con un aumento en la expresión de transportadores de colesterol en la membrana apical como NPC1L1 y CD36. La administración exógena de PCSK9 incrementó la secreción de QM al estimular la síntesis de lípidos y ApoB-4823.

Insulina. En condiciones de insulino-sensibilidad, esta hormona suprime la síntesis de QM y también de lipoproteínas de muy baja densidad (VLDL). Se demostró en sujetos sanos que la producción de QM se suprimió durante un clamp euglucémico hiperinsulinémico24. En este trabajo se observó que este efecto fue independiente de la acción de la insulina sobre los niveles de ácidos grasos libres plasmáticos, lo que sugirió un rol directo de esta hormona sobre la producción de QM. Este efecto directo ha sido demostrado tanto en estudios in vitro como en modelos animales en los cuales la insulina disminuyó la expresión del gen de la MTP25. Se debe considerar que además de la glucemia, existen múltiples factores que regulan la secreción de insulina. Entre estos, se puede citar a la dopamina que, a través de sus receptores en la célula beta, disminuye la secreción de insulina dependiente de glucosa26.
Péptido similar al glucagón tipo 1(GLP-1). En modelo del hámster dorado sirio, (modelo de gran similitud al modelo humano en términos de metabolismo de QM) se demostró que un agonista de GLP-1 disminuyó la secreción de QM asociado a la disminución de ácidos grasos libres27. Los trabajos clínicos que han utilizado fármacos inhibidores de la dipeptidil peptidasa tipo 4 a fin de potenciar los efectos del GLP-1 endógeno han mostrado superioridad en la reducción de los triglicéridos postprandiales en comparación con los triglicéridos en ayunas28.
En sujetos sanos, la administración de un análogo de GLP-1 como el exenatide mostró una reducción significativa de la producción de ApoB-48 independientemente de la secreción de insulina y del vaciamiento gástrico29.
Péptido similar al glucagón tipo 2 (GLP-2). El GLP-2 es co-secretado con el GLP-1. Cumple un papel en la integridad de la mucosa intestinal. Se observó que la administración de GLP-2 en hámsteres produjo un aumento de la lipemia postprandial mediante el incremento de la secreción de QM tanto in vivo como ex vivo30. Esto se debe a que el GLP-2 aumentó la expresión de CD36 glicosilado aumentando la absorción de ácidos grasos libres31.
Glucosa y fructosa. Las dietas ricas en sacarosa acentúan la hipertrigliceridemia debido a la sobreproducción tanto de triglicéridos (aumento en el tamaño de las partículas) como de ApoB (aumento del número de partículas), junto con un deterioro en su depuración. A su vez, el consumo crónico de fructosa aumenta la lipogénesis de novo, promueve la dislipidemia y aumenta la adiposidad visceral32. También se demostró que la administración de glucosa por vía endovenosa estimuló la producción de ApoB-48 sin modificar su catabolismo33.
El intestino: ¿un órgano blanco de la insulina?
Como se mencionó precedentemente, la insulina regula la síntesis de QM a nivel del enterocito disminuyendo su producción. Se debe recordar que fisiológicamente la insulina se une a su receptor y tras esta unión se produce la fosforilación de los residuos de tirosina del receptor y esto inicia una cascada de respuestas muy complejas, activando los llamados sustratos del receptor de insulina 1 y 2. Existen dos vías divergentes de señalización asociadas con la activación del receptor de insulina: la vía de la fosfatidilinositol-3-kinasa/Akt que media las acciones metabólicas y la vía de la proteína-quinasa activada por mitógenos que se asocia con la función de la insulina como reguladora de la expresión de diversos genes. Se demostró en un estudio realizado en hámsteres insulino-resistentes luego de una alimentación rica en fructosa que en el enterocito resistente a la insulina se produce una disminución en la fosforilación de los residuos tirosina en el receptor de insulina y los sustratos del receptor de insulina 1, en conjunto con un aumento de la proteína tirosina-fosfatasa34. Esto evidenció en el enterocito refractario a la acción de la insulina un aumento en la estabilidad de la ApoB-48 y un incremento en la síntesis y secreción de lipoproteínas. También se encontraron niveles elevados de las quinasas relacionadas con la fosforilación extracelular 1 y 2 que son miembros de la vía de las quinasas activadas por mitógenos implicada en el inicio de la secreción intestinal de ApoB-48.
Asimismo, se informó que la administración de factor de necrosis tumoral interfirió en la señalización normal de la insulina a nivel intestinal, disminuyendo la fosforilación en los residuos tirosina tanto de su receptor como de los sustratos del receptor de insulina 1 y Akt. Estos hallazgos permitieron inferir que la inflamación podría ser, en parte, la responsable de la resistencia a la insulina a nivel intestinal y, por lo tanto, constituir la base fisiopatológica de la dislipidemia postprandial que se observa en los estados de insulino-resistencia35.
Fisiopatología de la dislipidemia de origen intestinal en la diabetes mellitus tipo 2
Por mucho tiempo se ha considerado que la alteración en la depuración de QM explica el incremento en sus niveles plasmáticos observado en los pacientes con DM2. Estudios recientes han demostrado un aumento en la tasa de producción de QM en situaciones como la insulino-resistencia36 y la DM237, tanto en ayunas como en estado postprandial. Dentro de la fisiopatología de la DM2 se considera que existe una disminución en la secreción de GLP-138. En concordancia con este concepto, el efecto incretina se demostró en estudios de intervención con análogos de GLP-139 o tras la administración de inhibidores de la enzima dipeptidil peptidasa tipo 440 con una reducción de los niveles plasmáticos de QM.
En un metaanálisis realizado por nuestro grupo se observó que el tratamiento con análogos de GLP-1 y con fármacos inhibidores de la enzima dipeptidil peptidasa tipo 4 redujo los niveles de Apo-B48 y de triglicéridos postprandiales en pacientes con DM241.
En forma similar a lo que sucede con el hepatocito, el enterocito incrementa la producción de QM en respuesta a la elevación de ácidos grasos libres plasmáticos. Esto podría deberse a la estimulación de los ácidos grasos libres sobre la expresión de la apolipoproteína C-III, la cual, a su vez, estimula la síntesis de QM42. En un reciente trabajo de nuestro equipo se observó que pacientes con obesidad y DM2 presentaron un incremento en los niveles de apolipoproteína C-III, los cuales disminuyeron a los 6 meses de realizarse una cirugía de bypass o manga gástrica, asociado a una mejoría de la insulino-resistencia43.
Si bien no está totalmente dilucidado, se considera que la sobreproducción crónica por parte del intestino de ApoB-48 es secundaria al hiperinsulinismo e insulino-resistencia asociado con mayor ensamblaje y secreción de QM contribuyendo a la hiperlipemia posprandial44.
En sujetos insulino-resistentes se observó un aumento en la tasa de producción de QM que se correlacionó con la insulinemia en ayunas36. Hogue y colaboradores37 confirmaron estas observaciones y demostraron un incremento en la tasa de producción de QM asociado con un descenso en su catabolismo en sujetos con DM2. En este trabajo se encontró que la condición de diabético fue un predictor independiente de la tasa de producción de QM. En una publicación reciente encontramos una disminución del efecto fisiológico supresor de la insulina sobre la secreción de QM en sujetos con DM2, lo que explica la hiperlipemia posprandial en estos pacientes45.
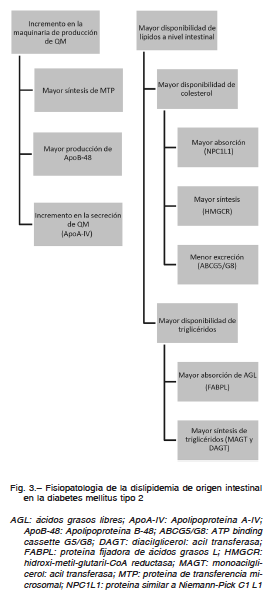
Se considera que el incremento en la producción de QM asociado con la DM2 (Fig. 3) puede deberse a:
1) Incremento en la maquinaria de producción de QM
2) Mayor disponibilidad de lípidos a nivel intracelular
1) Incremento en la maquinaria de producción de quilomicrón
En la síntesis de QM la MTP tiene un doble papel actuando tanto en la unión de la ApoB-48 con los triglicéridos como en la lipidación adicional del QM inmaduro.
Estudios realizados en sujetos con DM2 demostraron un aumento en la expresión intestinal de ARNm de la MTP46. Esta misma observación se confirmó en estudios realizados en ratas con DM247. A su vez, se demostró que la MTP juega un papel crucial en la regulación del contenido de colesterol en la partícula del QM dado que se encontró una relación positiva entre el contenido de ARNm de MTP y la fracción de colesterol en el QM en sujetos con DM248.
Se observó en un estudio realizado por Zoltowska y colaboradores49 mediante el análisis ex vivo de yeyuno del roedor Psammomys obesus con DM2 un aumento en la biogénesis de la ApoB-48 asociado con una reducción de su degradación proteosomal.
La apolipoproteína A-IV interviene en el transporte intracelular de QM y estimula la liberación de triglicéridos por la membrana basolateral incrementando el tamaño de las lipoproteínas secretadas14. Se comprobó un incremento en la expresión de ARNm de la apolipoproteína A-IV en biopsias de yeyuno de sujetos obesos mórbidos con DM250.
2) Mayor disponibilidad de lípidos a nivel intracelular
Si bien el QM representa una lipoproteína rica en triglicéridos, se requiere la disponibilidad tanto de triglicéridos como colesterol para su síntesis.
En lo que respecta a la mayor disponibilidad de colesterol, se demostró en pacientes con DM2 un aumento en la absorción de colesterol asociada con un incremento en el número de transportadores NPC1L146. Asimismo, Ravid y colaboradores51 demostraron que la hiperglucemia aumentó la absorción de colesterol por aumento en la expresión de NPC1L1. Se observó un resultado similar en células CaCo2 donde se descubrió que retirando la glucosa del medio de cultivo disminuyó la expresión de ARNm de NPC1L1. A su vez, en dicha investigación la glucosa promovió la actividad de NPC1L1 en una forma dependiente de su concentración52. Este incremento en los niveles de NPC1L1 en presencia de DM2 o hiperglucemia permitiría el uso de fármacos que bloqueen la absorción del colesterol.
Además, se demostró un aumento en la expresión de ARNm de la enzima limitante en la síntesis de colesterol, la hidroxi-metil-glutaril-CoA reductasa, en un experimento realizado en ratas con DM247. Sumado a esto, se comprobó en sujetos con DM2 una concentración de ARNm significativamente menor del heterodímero ABCG5/8 asociado a menor excreción del colesterol desde el enterocito hacia la luz intestinal. Se observó una correlación negativa entre los niveles de ARNm de la MTP y los niveles de ARNm de ABCG5/G8 sugiriendo que un incremento en los niveles disponibles de colesterol podría estimular la expresión de MTP y conducir a un incremento significativo en el número de partículas de QM46.
Con respecto a los ácidos grasos, también se observó en el estudio Zoltowska y colaboradores49 un incremento en la actividad de las enzimas mono y diacilglicerol: acil transferasa, enzimas clave en la síntesis de triglicéridos. A su vez, se observó en dicha investigación un incremento en la proteína fijadora de ácidos grasos L, lo cual sugiere un incremento en el transporte intracelular de lípidos en animales con DM2.
En los estados de resistencia a la insulina y DM2 se observa un aumento en la concentración de ácidos grasos libres plasmáticos debido al incremento en la lipólisis en el tejido adiposo por la menor acción de la insulina sobre la lipasa hormono sensible. Esto origina que esos ácidos grasos sean re-direccionados hacia tejidos entre los que se incluyen el intestino, el hígado y el músculo.
Además, la disminución plasmática de GLP-1 se asoció con un aumento del transporte de ácidos grasos libres en la membrana apical del enterocito por intermedio de un mayor transporte del CD36 en modelos de resistencia a la insulina y DM253.
A modo de síntesis, se puede observar que en presencia de DM2, el enterocito cuenta con una mayor disponibilidad de lípidos y se demostró que en dicha situación las partículas de QM poseen una cantidad significativamente mayor tanto de triglicéridos como de colesterol47, 48.
Rol del quilomicrón en la aterosclerosis
El mecanismo por el cual la dislipidemia típica de la DM2 acelera el proceso aterosclerótico es multifactorial y complejo. Se considera actualmente que el enterocito juega un papel clave en la dislipidemia asociada con los estados de resistencia a la insulina54.
Como se mencionó previamente, la concentración de triglicéridos en estado postprandial es un predictor fuerte de riesgo cardiovascular, independiente de los factores de riesgo tradicionales. El aumento del riesgo cardiovascular asociado a la hiperlipemia postprandial se atribuiría a las propiedades inflamatorias y aterogénicas del QM generadas en el estado postprandial. El QM puede actuar directamente en el proceso de aterosclerosis dado que los ácidos grasos libres circulantes constituyen ligandos endógenos de los receptores tipo Toll presente en los leucocitos. De esta forma activan al factor de transcripción nuclear Kappa B y estimulan la síntesis de citoquinas y generan un microambiente proinflamatorio, procoagulante y proaterogénico en la pared arterial55.
Los remanentes de QM parcialmente catabolizados también tendrían propiedades proaterogénicas. Se demostró la presencia de partículas de ApoB-48 en remanentes de QM de placas de aterosclerosis. Se identificó un sitio de unión para la ApoB-48 en los proteoglicanos de la íntima arterial56. A su vez, se aisló en macrófagos y monocitos un receptor específico para la ApoB-48. Se informó la presencia de dicho receptor en las células espumosas de las estrías lipídicas y en las placas de ateroma. Es probable que su capacidad de enlace y de internalización de remanentes de QM participe en la adquisición del fenotipo foam cell de los macrófagos y en la constitución de las lesiones de aterosclerosis55.
Teniendo en cuenta su concentración plasmática, la capacidad de retención de los remanentes de VLDL con ApoB-100 es 10 veces superior en relación a los remanentes de QM. Sin embargo, el contenido de colesterol es 40 veces superior en el QM en comparación con las VLDL con ApoB-100. Esta condición otorga al remanente de QM la posibilidad de liberar 4 veces más colesterol en la pared vascular que las VLDL con ApoB-10057. En consecuencia, la captación de remanentes de QM a nivel de las arterias en el período postprandial podría aportar un riesgo aterogénico importante. Quizás este riesgo sea aún mayor en condiciones en las cuales su concentración plasmática postprandial se encuentra aumentada.
En conclusión, en la DM2 el enterocito presenta una mayor disponibilidad de substratos y de proteínas involucradas en la síntesis y secreción de QM. Esto se suma a la alteración del catabolismo de estas lipoproteínas que presentan las personas con DM2. La administración de fármacos que potencian la acción del GLP-1 reduce la concentración de QM en pacientes con DM2.
La hiperlipemia postprandial representa un importante factor de riesgo cardiovascular residual en sujetos con DM2. Teniendo en cuenta la capacidad aterogénica de los remanentes de QM, se requieren investigaciones adicionales para la comprensión integral de los mecanismos moleculares que contribuyen a la disfunción del enterocito y la sobreproducción de QM presentes en la DM2. Esto podría tener implicancias tanto en el seguimiento de los pacientes como para futuras dianas terapéuticas.
Conflicto de intereses: Ninguno para declarar
Bibliografía
1. Hokanson JE, Austin MA. Plasma triglyceride level is a risk factor for cardiovascular disease independent of high-density lipoprotein cholesterol level: a meta-analysis of population-based prospective studies. J Cardiovasc Risk 1996; 3: 213-9.
2. Nordestgaard BG, Benn M, Schnohr P, Tybjaerg-Hansen A. Nonfasting triglycerides and risk of myocardial infarction, ischemic heart disease, and death in men and women. JAMA 2007; 298: 299-308.
3. Nakamura K, Miyoshi T, Yunoki K, Ito H. Postprandial hyperlipidemia as a potential residual risk factor. J Cardiol 2016; 67: 335-9.
4. Zhang J-H, Ge L, Qi W, et al. The N-terminal domain of NPC1L1 protein binds cholesterol and plays essential roles in cholesterol uptake. J Biol Chem 2011; 286: 25088-97.
5. Duan L-P, Wang HH, Wang DQ-H. Cholesterol absorption is mainly regulated by the jejunal and ileal ATP-binding cassette sterol efflux transporters Abcg5 and Abcg8 in mice. J Lipid Res 2004; 45: 1312-23.
6. Blanchard C, Moreau F, Cariou B, Le May C. Trans-intestinal cholesterol excretion (TICE): a new route for cholesterol excretion. Med Sci 2014; 30: 896-901.
7. Van der Veen JN, Van Dijk TH, Vrins CL, et al. Activation of the liver X receptor stimulates trans-intestinal excretion of plasma cholesterol. J Biol Chem 2009; 284: 19211-9.
8. Nassir F, Wilson B, Han X, Gross RW, Abumrad NA. CD36 is important for fatty acid and cholesterol uptake by the proximal but not distal intestine. J Biol Chem 2007; 282: 19493-501.
9. Milger K, Herrmann T, Becker C, et al. Cellular uptake of fatty acids driven by the ER-localized acyl-CoA synthetase FATP4. J Cell Sci 2006; 119: 4678-88.
10. Pan X, Hussain MM. Gut triglyceride production. Biochim Biophys Acta 2012; 1821: 727-35.
11. Hagan DL, Kienzle B, Jamil H, Hariharan N. Transcriptional regulation of human and hamster microsomal triglyceride transfer protein genes. Cell type-specific expression and response to metabolic regulators. J Biol Chem 1994; 269: 28737-44.
12. Sato R, Miyamoto W, Inoue J, Terada T, Imanaka T, Maeda M. Sterol regulatory element-binding protein negatively regulates microsomal triglyceride transfer protein gene transcription. J Biol Chem 1999; 274: 24714-20.
13. Jattan J, Rodia C, Li D, et al. Using primary murine intestinal enteroids to study dietary TAG absorption, lipoprotein synthesis, and the role of apoC-III in the intestine. J Lipid Res 2017; 58: 853-65.
14. Kohan AB, Wang F, Lo CM, Liu M, Tso P. ApoA-IV: current and emerging roles in intestinal lipid metabolism, glucose homeostasis, and satiety. Am J Physiol Gastrointest Liver Physiol 2015; 308: G472-81.
15. Davidson NO, Shelness GS. Apolipoprotein B: mRNA editing, lipoprotein assembly, and presecretory degradation. Annu Rev Nutr 2000; 20: 169-93.
16. Karpe F, Steiner G, Uffelman K, Olivecrona T, Hamsten A. Postprandial lipoproteins and progression of coronary atherosclerosis. Atherosclerosis 1994; 106: 83-97.
17. Morita SY. Metabolism and Modification of Apolipoprotein B-Containing Lipoproteins Involved in Dyslipidemia and Atherosclerosis. Biol Pharm Bull 2016; 39: 1-24.
18. Xiao C, Lewis GF. Regulation of chylomicron production in humans. Biochim Biophys Acta 2012; 1821: 736-46.
19. Tran TT, Poirier H, Clément L, et al. Luminal lipid regulates CD36 levels and downstream signaling to stimulate chylomicron synthesis. J Biol Chem 2011; 286: 25201-10.
20. Duez H, Lamarche B, Valéro R, et al. Both intestinal and hepatic lipoprotein production are stimulated by an acute elevation of plasma free fatty acids in humans. Circulation 2008; 117: 2369-76.
21. Lewis GF, Naples M, Uffelman K, Leung N, Szeto L, Adeli K. Intestinal lipoprotein production is stimulated by an acute elevation of plasma free fatty acids in the fasting state: studies in insulin-resistant and insulin-sensitized Syrian golden hamsters. Endocrinology 2004; 145: 5006-12.
22. Iqbal J, Rudel LL, Hussain MM. Microsomal triglyceride transfer protein enhances cellular cholesteryl esterification by relieving product inhibition. J Biol Chem 2008; 283: 19967-80.
23. Rashid S, Tavori H, Brown PE, et al. Proprotein convertase subtilisin kexin type 9 promotes intestinal overproduction of triglyceride-rich apolipoprotein B lipoproteins through both low-density lipoprotein receptor-dependent and -independent mechanisms. Circulation 2014; 130: 431-41.
24. Pavlic M, Xiao C, Szeto L, Patterson BW, Lewis GF. Insulin acutely inhibits intestinal lipoprotein secretion in humans in part by suppressing plasma free fatty acids. Diabetes 2010; 59: 580-7.
25. van Greevenbroek MM, de Bruin TW. Chylomicron synthesis by intestinal cells in vitro and in vivo. Atherosclerosis 1998; 141: 9-16.
26. Lopez Vicchi F, Luquea G, Briea B, Nogueira JP, Garcia Tornadua I, Becu-Villalobos D. Dopaminergic drugs in type 2 diabetes and glucose homeostasis. Pharmacol Res 2016; 109: 74-80.
27. Hsieh J, Longuet C, Baker CL, et al. The glucagon-like peptide 1 receptor is essential for postprandial lipoprotein synthesis and secretion in hamsters and mice. Diabetologia 2010; 53: 552-61.
28. Xiao C, Dash S, Morgantini C, Patterson BW, Lewis GF. Sitagliptin, a DPP-4 inhibitor, acutely inhibits intestinal lipoprotein particle secretion in healthy humans. Diabetes 2014; 63: 2394-401.
29. Xiao C, Bandsma RHJ, Dash S, Szeto L, Lewis GF. Exenatide, a glucagon-like peptide-1 receptor agonist, acutely inhibits intestinal lipoprotein production in healthy humans. Arterioscler Thromb Vasc Biol 2012; 32: 1513-9.
30. Hein GJ, Baker C, Hsieh J, Farr S, Adeli K. GLP-1 and GLP-2 as yin and yang of intestinal lipoprotein production: evidence for predominance of GLP-2-stimulated postprandial lipemia in normal and insulin-resistant states. Diabetes 2013; 62: 373-81.
31. Hsieh J, Longuet C, Maida A, et al. Glucagon-like peptide-2 increases intestinal lipid absorption and chylomicron production via CD36. Gastroenterology 2009; 137: 997-1005.
32. Stanhope KL, Schwarz JM, Keim NL, et al. Consuming fructose-sweetened, not glucose-sweetened, beverages increases visceral adiposity and lipids and decreases insulin sensitivity in overweight/obese humans. J Clin Invest 2009; 119: 1322-34.
33. Xiao C, Dash S, Morgantini C, Lewis GF. Intravenous Glucose Acutely Stimulates Intestinal Lipoprotein Secretion in Healthy Humans. Arterioscler Thromb Vasc Biol 2016; 36: 1457-63.
34. Federico LM, Naples M, Taylor D, Adeli K. Intestinal insulin resistance and aberrant production of apolipoprotein B48 lipoproteins in an animal model of insulin resistance and metabolic dyslipidemia: evidence for activation of protein tyrosine phosphatase-1B, extracellular signal-related kinase, and sterol regulatory element-binding protein-1c in the fructose-fed hamster intestine. Diabetes 2006; 55: 1316-26.
35. Hsieh J, Hayashi AA, Webb J, Adeli K. Postprandial dyslipidemia in insulin resistance: mechanisms and role of intestinal insulin sensitivity. Atheroscler Suppl 2008; 9: 7-13.
36. Duez H, Lamarche B, Uffelman KD, Valero R, Cohn JS, Lewis GF. Hyperinsulinemia is associated with increased production rate of intestinal apolipoprotein B-48-containing lipoproteins in humans. Arterioscler Thromb Vasc Biol 2006; 26: 1357-63.
37. Hogue JC, Lamarche B, Tremblay AJ, Bergeron J, Gagné C, Couture P. Evidence of increased secretion of apolipoprotein B-48-containing lipoproteins in subjects with type 2 diabetes. J Lipid Res 2007; 48: 1336-42.
38. DeFronzo RA. From the triumvirate to the ominous octet: A new paradigm for the treatment of type 2 diabetes mellitus. Diabetes 2009; 58: 773-95.
39. Schwartz EA, Koska J, Mullin MP, Syoufi I, Schwenke DC, Reaven PD. Exenatide suppresses postprandial elevations in lipids and lipoproteins in individuals with impaired glucose tolerance and recent onset type 2 diabetes mellitus. Atherosclerosis 2010; 212: 217-22.
40. Matikainen N, Mänttäri S, Schweizer A, et al. Vildagliptin therapy reduces postprandial intestinal triglyceride-rich lipoprotein particles in patients with type 2 diabetes. Diabetologia 2006; 49: 2049-57.
41. Cartazzo A, Nogueira JP. Meta-análisis del efecto incretina sobre el metabolismo de lipoproteínas ricas en triglicéridos intestinales en diabetes tipo 2. Rev Arg de Lipidos 2017; 1: 42-6.
42. Pavlic M, Valéro R, Duez H, et al. Triglyceride-rich lipoprotein-associated apolipoprotein C-III production is stimulated by plasma free fatty acids in humans. Arterioscler Thromb Vasc Biol 2008; 28: 1660-5.
43. Maraninchi M, Padilla N, Béliard S, et al. Impact of bariatric surgery on apolipoprotein C-III levels and lipoprotein distribution in obese human subjects. J Clin Lipidol 2017; 11: 495-506.
44. Duez H, Pavlic M, Lewis GF. Mechanism of intestinal lipoprotein overproduction in insulin resistant humans. Atheroscler Suppl 2008; 9: 33-8.
45. Nogueira J-P, Maraninchi M, Béliard S, et al. Absence of acute inhibitory effect of insulin on chylomicron production in type 2 diabetes. Arterioscler Thromb Vasc Biol 2012; 32: 1039-44.
46. Lally S, Tan CY, Owens D, Tomkin GH. Messenger RNA levels of genes involved in dysregulation of postprandial lipoproteins in type 2 diabetes: the role of Niemann-Pick C1-like 1, ATP-binding cassette, transporters G5 and G8, and of microsomal triglyceride transfer protein. Diabetologia 2006; 49: 1008-16.
47. Lally S, Owens D, Tomkin GH. The different effect of pioglitazone as compared to insulin on expression of hepatic and intestinal genes regulating post-prandial lipoproteins in diabetes. Atherosclerosis 2007; 193: 343-51.
48. Phillips C, Mullan K, Owens D, Tomkin GH. Intestinal microsomal triglyceride transfer protein in type 2 diabetic and non-diabetic subjects: the relationship to triglyceride-rich postprandial lipoprotein composition. Atherosclerosis 2006; 187: 57-64.
49. Zoltowska M, Ziv E, Delvin E, et al. Cellular aspects of intestinal lipoprotein assembly in Psammomys obesus: a model of insulin resistance and type 2 diabetes. Diabetes 2003; 52: 2539-45.
50. Soriguer F, García-Serrano S, Garrido-Sánchez L, et al. Jejunal wall triglyceride concentration of morbidly obese persons is lower in those with type 2 diabetes mellitus. J Lipid Res 2010; 51: 3516-23.
51. Ravid Z, Bendayan M, Delvin E, et al. Modulation of intestinal cholesterol absorption by high glucose levels: impact on cholesterol transporters, regulatory enzymes, and transcription factors. Am J Physiol Gastrointest Liver Physiol 2008; 295: 873-85.
52. Malhotra P, Boddy CS, Soni V, et al. D-Glucose modulates intestinal Niemann-Pick C1-like 1 (NPC1L1) gene expression via transcriptional regulation. Am J Physiol Gastrointest Liver Physiol 2013; 304: 203-10.
53. Xiao C, Dash S, Lewis GF. Mechanisms of incretin effects on plasma lipids and implications for the cardiovascular system. Cardiovasc Hematol Agents Med Chem 2012; 10: 289-94.
54. Nogueira JP, Brites FD. Role of enterocytes in dyslipidemia of insulin-resistant states. Endocrinol Nutr 2013; 60: 179-89.
55. Alipour A, Elte JWF, van Zaanen HCT, Rietveld AP, Castro Cabezas M. Novel aspects of postprandial lipemia in relation to atherosclerosis. Atheroscler Suppl 2008; 9: 39-44.
56. Flood C, Gustafsson M, Richardson PE, Harvey SC, Segrest JP, Borén J. Identification of the proteoglycan binding site in apolipoprotein B48. J Biol Chem 2002; 277: 32228-33.
57. Proctor SD, Vine DF, Mamo JCL. Arterial retention of apolipoprotein B(48)- and B(100)- containing lipoproteins in atherogenesis. Curr Opin Lipidol 2002; 13: 461-70.
– – – –
Sobre la claridad he de deciros que debe ser vuestra más vehemente aspiración. El solo intento de sacar al sol la propia tiniebla es ya plausible. Luego, como dicen en Aragón: ¡Veremos!
Antonio Machado (1875-1939)
Juan de Mairena II. Buenos Aires: Losada, 1968. 4ta. Edición, p 30