
SANDRA G. VLACHOVSKY 1, DAIANA S. SÁNCHEZ 2, LUIS A. DI CIANO 1, ELISABET M. ODDO 1, PABLO J. AZURMENDI 1, CLAUDIA SILBERSTEIN 2, FERNANDO R. IBARRA 1, 2
1 Laboratorio de Nefrología Experimental y Bioquímica Molecular, Instituto de Investigaciones Médicas A. Lanari, Universidad de Buenos Aires, 2 Departamento de Ciencias Fisiológicas, Instituto de Fisiología y Biofísica Bernardo Houssay (IFIBIO Houssay)- CONICET, Facultad de Medicina, Universidad de Buenos Aires, Argentina
Resumen Un tercio de la población mundial tiene niveles anormalmente altos de presión arterial, hipertensión, responsable de casi el 50% de las muertes por accidente cerebrovascular y enfermedad coronaria. La sensibilidad a la sal es un factor de riesgo para la morbilidad y mortalidad cardiovascular y también para otras enfermedades. En estudios previos describimos un modelo de hipertensión sal sensible (HSS) en ratas Wistar ovariectomizadas (oVx) adultas. Las ratas oVx son normotensas con ingesta normal de sal (NS, 0.24% de NaCl), pero desarrollan un perfil de HSS con una ingesta elevada de sal (HS, 1% de NaCl). En los estudios en riñón encontramos que el circuito receptor D1 de dopamina, citocromo P450 4A y Na+, K+-ATPasa está alterado por la ausencia de hormonas ováricas, lo que se asocia a menor excreción de sodio e hipertensión arterial. La ingesta HS en ratas oVx también promueve cambios en la expresión de proteínas relacionadas con el transporte de sodio en células mononucleares de sangre periférica, principalmente linfocitos periféricos. Por lo tanto, el transporte de sodio se modifica en varios niveles de la fisiología normal. En estudios recientes observamos que el estradiol aumenta la proliferación y diferenciación de células epiteliales en cultivos de corteza renal humana. Sensibilidad a la sal, inmunidad adaptativa, presión arterial y proliferación de células epiteliales en riñón son fenómenos de gran importancia biológica regulados por estradiol.
Palabras clave: hormonas femeninas, función renal, hipertensión sensible a la sal, inmunidad adaptativa, proliferación epitelial
Abstract Female sex hormones participate in the regulation of blood pressure and renal epithelial proliferation, effects not related to their reproductive function. About one-third of the world’s population has abnormally high levels of blood pressure, hypertension, which is responsible for almost 50% of deaths from stroke and coronary heart disease. Salt sensitivity is a risk factor for cardiovascular morbidity and mortality and other diseases as well. We reported a model of salt sensitive hypertension in adult ovariectomized (oVx) Wistar rats. oVx rats are normotensive under normal salt intake (NS, 0.24% NaCl), but upon a high salt intake (HS, 1% NaCl) oVx rats developed a blood pressure profile of salt-sensitive hypertension. Our studies on kidney molecules related to sodium balance found that the circuit dopamine D1-like receptor, cytochrome P450 4A and Na+, K+-ATPase is altered by the absence of ovary hormones which is accompanied by a reduced ability to excrete sodium. In oVx rats HS intake also promotes changes in the expression of proteins related to sodium transport in peripheral blood mononuclear cells, mainly peripheral lymphocytes. Therefore, sodium transport is modified at several levels of normal physiology. Lately, we described that estradiol increases the rate of renal epithelial cell proliferation in primary cultures developed from human renal cortex. Thus, salt sensitivity, adaptive immunity, blood pressure and renal cell proliferation are complex biological responses regulated by female sex hormones.
Key words: female hormones, renal function, salt-sensitive hypertension, adaptive immunity, renal cell proliferation
Dirección postal: Sandra G. Vlachovsky, Laboratorio de Nefrología Experimental y Bioquímica Molecular, Instituto de Investigaciones Médicas A. Lanari, Combatientes de Malvinas 3150, 1427 Buenos Aires, Argentina
e-mail: sandragvla@gmail.com
Las hormonas sexuales femeninas tienen múltiples efectos sistémicos y órgano-específicos no vinculados a su función reproductiva. En este artículo describimos experiencias de diferentes grupos de investigación y propias, sobre aspectos vinculados a la regulación de la presión arterial (PA) y de la proliferación epitelial renal. En cuanto a la PA, se destaca el desarrollo de hipertensión arterial (HTA) sensible a la sal y de un componente inmunológico vinculado a la inmunidad adaptativa. El fenómeno de los cambios de PA, particularmente el incremento de la PA al aumentar la ingesta de sodio, ha despertado el interés de los investigadores desde tiempo atrás 1. Si bien en este artículo resaltamos la importancia de la ingesta excesiva de sodio, también hay que tener en cuenta que frecuentemente se acompaña de otros hábitos alimentarios y de actividad física inadecuados como consumo excesivo de hidratos de carbono y sedentarismo. La combinación de estos factores lleva al desarrollo de síndrome metabólico 2.
La regulación de la proliferación epitelial renal podría ser muy importante en la reparación del epitelio dañado. En trabajos anteriores describimos que el crecimiento renal compensador luego de nefrectomía unilateral en ratas intactas o gonadectomizadas es diferente en ratas hembras y machos. El mayor crecimiento e hipertrofia se observó en machos intactos. Estos hallazgos resaltan la importancia del ambiente hormonal en cada caso 3.
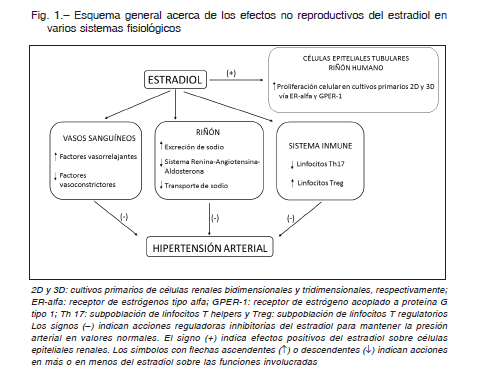
En la Figura 1 se describen estos mecanismos no vinculados a la reproducción de las hormonas sexuales
femeninas.
Hormonas sexuales e hipertensión sensible a la sal
El perfil de sensibilidad a la sal o hipertensión sensible a la sal (HSS), se observa cuando al aumentar la ingesta de NaCl o sal de mesa, aumenta la PA. Este es el concepto predominante pero también suele definirse como un descenso de la PA al reducir la ingesta de NaCl. Las personas más afectadas son las de edad avanzada, ya que un 80% de los hipertensos mayores de 60 años padecen HSS, mientras que solo el 50% de los menores de 40 años la presentan 4. Aunque la sensibilidad a la sal no es una respuesta observada en un gran porcentaje de la población o de los animales de experimentación, en lo que llamaríamos condiciones basales o habituales de vida 5, se la asocia con el aumento de morbi-mortalidad cardiovascular (CV) incluyendo enfermedad renal 2, como ocurre en otros cuadros de HTA.
Un rasgo característico también sería que, una vez identificado este patrón de respuesta CV, podría evitarse disminuyendo la ingesta de NaCl. En la práctica clínica, esto ocurre tanto en pacientes con HSS como en pacientes con HTA con otras etiologías pero que mejoran al disminuir la ingesta de NaCl. Además, es interesante señalar que una población aún indeterminada de las personas clasificadas dentro de los rangos de normotensión, también presentan el fenómeno de sensibilidad a la sal: su PA se modifica más que en otros normotensos al cambiar la ingesta de sal, sin llegar a la HTA. Este patrón de respuesta se asocia también a mayor prevalencia de enfermedad CV y a otras enfermedades como neoplasias 1.
Cuando se estudian grupos comparables de mujeres y hombres, incluso animales de experimentación hembras y machos, se observa que la PA es consistentemente menor en mujeres/hembras en edad reproductiva que en hombres/machos 6, 7. En estudios en animales de experimentación se aprecia que las hembras pueden excretar una sobrecarga de sodio con mayor rapidez y eficacia que los machos 8, 9. Junto con esto, los eventos CV también son menores en mujeres que en hombres, excepto que concurran otros factores como tabaquismo, obesidad o consumo de drogas. Además, se observa que de producirse enfermedad o lesión renal las mujeres en edad reproductiva presentan una evolución mejor y más lenta que los hombres 10. En el mismo sentido, estudios experimentales de injuria o lesión renal aguda también señalan un patrón de recuperación o deterioro de la función renal de acuerdo a la edad y sexo, siendo las ratas hembras jóvenes las que resisten mejor la lesión aguda y se recuperan con mayor prontitud 11.
Este perfil cambia en la postmenopausia. Una década después de la menopausia el patrón de protección CV y renal disminuye y se aproxima en su respuesta biológica o sistémica a su contrapartida masculina 12, 13.
Es frecuente que las mujeres postmenopáusicas desarrollen HTA con un patrón de respuesta con sensibilidad a la sal 13. Este cambio del perfil de respuesta CV y a la ingesta de sodio se asocia al descenso de los niveles de hormonas ováricas, fundamentalmente estrógenos 10, un rasgo también descripto en varios modelos de animales de experimentación 7.
Diversos grupos de investigadores han centrado los estudios en los aspectos vasculares de la HTA que se presentan en mujeres postmenopáusicas: los estrógenos favorecen la síntesis de óxido nítrico vascular 14 mientras que su ausencia produce su disminución con un aumento concomitante de la respuesta a la angiotensina II seguido de vasoconstricción e HTA 15-17.
Hormonas sexuales femeninas y regulación del sodio en riñón
Los estudios clásicos experimentales se hicieron con animales machos o con hembras que recibían mineralocorticoides y sal (DOCA-Sal) evaluando sus efectos sistémicos18 o con infusión crónica de angiotensina II 19-21.
Dichos estudios tradicionales relacionan la HTA con factores que retienen sodio o incrementan el tono vascular de las arterias, como la angiotensina y la noradrenalina.
Esta hipótesis inicial se ha ido modificando con los años debido a la gran cantidad de estudios que han ampliado los conocimientos sobre este tema mostrando que la baja disponibilidad de factores natriuréticos y vasodilatadores pueden predisponer a la HTA. Varias líneas de evidencia sugieren que la dopamina renal, entre otros factores vasodilatadores y natriuréticos como factor natriurético atrial, óxido nítrico, prostaglandinas o bradiquinina, juega un papel preponderante en la fisiopatología de la HTA 22.
En este sentido, nuestro grupo ha desarrollado una serie de estudios en un modelo experimental en ratas Wistar hembras adultas ovariectomizadas para evaluar el perfil de respuesta de la PA y los transportadores o mediadores vinculados al balance de sodio en el riñón y su comportamiento ante un ingreso mayor de NaCl. En las primeras investigaciones encontramos que ante una ingesta normal de NaCl la actividad del sistema de calicreína-quinina y la expresión del gen de calicreína están aumentadas en tejido renal. En este modelo animal, la PA se mantiene normal o incluso disminuye 23.
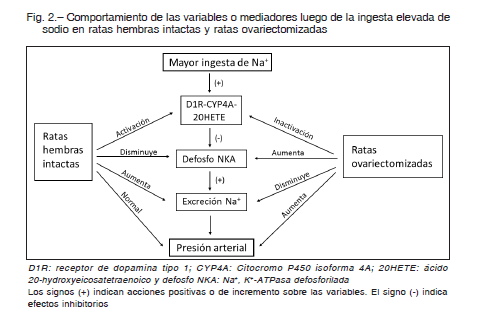
Posteriormente, analizamos la respuesta de la PA y de los transportadores de sodio en el riñón, comparando una situación basal con ingesta normal de sodio y otra con aumento de la ingesta de NaCl cuatro veces mayor 24. Ante la alta ingesta, las ratas ovariectomizadas excretan menos sodio que las intactas y desarrollan HSS. En cuanto a la función renal, se observó que en dichas ratas la expresión y función del receptor D1 de dopamina se encuentran disminuidas.
Esto se asocia con una expresión aumentada de la bomba Na+, K+-ATPasa (NKA). Es conocido que la NKA se regula por mecanismos de fosforilación/defosforilación en los que la dopamina es uno de sus moduladores y que, al ingerir más sodio, la dopamina renal incrementa su actividad para fosforilar a la NKA e inactivarla y, como efecto final, producir natriuresis. Nuestros resultados muestran que este mecanismo postraduccional se altera en las ratas ovariectomizadas. A ello se le suma que el citocromo P450 4A, uno de los mediadores moleculares por los que actúa la dopamina y que inhibe la NKA fosforilándola 25, se encuentra disminuido en su síntesis y actividad, lo que explicaría la defosforilación y aumento de actividad de NKA lo que llevaría, en consecuencia, a la retención de sodio.
De esta manera, se evidencia que las hormonas ováricas desempeñan una función clave en la regulación de la función renal y de la presión arterial. En la Figura 2 se esquematizan los hallazgos vinculados a una ingesta elevada de sodio comparando entre las ratas intactas y las ovariectomizadas.
Hormonas sexuales femeninas y sistema inmune
En modelos de HTA se han hallado alteraciones de la inmunidad adaptativa y en particular de los linfocitos T. Guzik y colaboradores 26 fueron los primeros en mostrar en ratas macho que los linfocitos T participan de la respuesta hipertensiva a la infusión crónica de angiotensina II. Posteriormente, se observó que en ratas hembra el modelo planteado no se repitió como en machos 27. Es decir, que la respuesta de las células T es específica del sexo en el que se estudie, resaltando la importancia de las hormonas femeninas. Como se observa en la Figura 1, el estradiol disminuye la diferenciación de linfocitos T hacia subpoblaciones proinflamatorias Th17 e incrementa los linfocitos reguladores o Treg antiinflamatorios 28, 29.
En estudios recientes, pudimos observar que las alteraciones en los receptores y transportadores relacionados al sodio en las ratas oVx también se aprecian en células mononucleares de sangre periférica, que en la rata predominantemente son linfocitos 30.
Estrógenos y proliferación celular del epitelio tubular
Otra función sistémica relacionada con las hormonas femeninas, en particular con los estrógenos, es que pueden estar involucrados en la regulación de la proliferación celular en tejidos renales (Fig. 1). La mayoría de las células de los túbulos renales se mantienen en estado quiescente, sin embargo, pueden adquirir la capacidad de proliferar ante un estímulo o daño 31, 32. En estudios de cultivos primarios de células epiteliales de corteza renal humana provenientes de riñones pediátricos, demostramos que el estradiol tiene la capacidad de inducir proliferación celular epitelial renal 33. Además, hemos desarrollado un cultivo primario tridimensional (cultivos 3D) de células epiteliales renales humanas que al proliferar sobre una matriz especial desarrollan túbulos renales de estructura comparable a la de un túbulo proximal nativo y también modifican su diferenciación y proliferación ante una injuria 34. El estradiol aumenta la capacidad proliferativa y la velocidad de formación de dichas estructuras tubulares 33. También se observó que estas células expresan tanto los receptores clásicos de estradiol ER-alfa y ER-beta como G proteincoupled estrogen receptor-1(GPER-1). Se observó que el estímulo de estradiol sobre la proliferación de las células epiteliales depende de la activación de ER-alfa y GPER-1, pero no de ER-beta 33.
Resumiendo, y en concordancia con investigaciones de otros grupos, mostramos por medio de experimentos sistémicos en animales y de cultivos primarios de células humanas, que las hormonas femeninas cumplen un papel importante en la regulación de la función renal y de la presión arterial.
Perspectivas: En estos efectos se hallan también involucrados mecanismos vinculados a la inmunidad adaptativa de linfocitos periféricos y fenómenos regulatorios de la proliferación tubular renal. Estos hallazgos muestran la importancia fundamentalmente del estradiol en estos efectos y la probabilidad de poder explorar o aplicar en el humano dichas investigaciones en el futuro.
Agradecimientos: Se agradece la asistencia técnica de Jorge E. Toledo en la ejecución de los trabajos de investigación que aquí se mencionan. Se agradece a la Secretaría de Ciencia y Técnica del Rectorado de la Universidad de Buenos Aires que financió las investigaciones que se mencionan mediante los proyectos UBACYT 20020120100379BA y 20020160100078BA
Conflicto de intereses: Ninguno para declarar
Bibliografía
1. de Wardener HE, MacGregor GA. Harmful effects of dietary salt in addition to hypertension. J Hum Hypertens 2002; 16: 213-23.
2. Global Status Report on noncommunicable diseases 2010. World Health Organization 2011. En: http://whqlibdoc.who.int/publications; consultado julio 2019.
3. Azurmendi PJ, Oddo EM, Toledo JE, Martin RS, Ibarra FR, Arrizurieta EE. Sexual hormones modulate compensatory renal growth and function. Medicina (B Aires) 2013; 73: 513-9.
4. Weinberger MH, Fineberg NS, Fineberg SE, Weinberger M. Salt sensitivity, pulse pressure, and death in normal and hypertensive humans. Hypertension 2001; 37: 429-32.
5. Bie P. Mechanisms of sodium balance: total body sodium, surrogate variables, and renal sodium excretion. Am J Physiol Regul Integr Comp Physiol 2018; 315: R945-62.
6. Maranon R, Reckelhoff JF. Sex and gender differences in control of blood pressure. Clin Sci (Lond) 2013; 125: 311-8.
7. Reckelhoff JF. Gender differences in the regulation of blood pressure. Hypertension 2001; 37: 1199-208.
8. Veiras LC, Girardi ACC, Curry J, et al. Sexual dimorphic pattern of renal transporters and electrolyte homeostasis. J Am Soc Nephrol 2017; 28: 3504-17.
9. Di Ciano LA, Azurmendi PJ, Vlachovsky SG, et al. Diferencias de género en presión arterial, función renal y respuesta a la dieta hipersódica en ratas Wistar. Rev Nefrol Dial Traspl 2018; 1: 15-7.
10. Pechere-Bertschi A, Burnier M. Gonadal steroids, salt-sensitivity and renal function. Curr Opin Nephrol Hypertens 2007; 16: 16-21.
11. Boddu R, Fan C, Rangarajan S, Sunil B, Bolisetty S, Curtis LM. Unique sex- and age-dependent effects in protective pathways in acute kidney injury. Am J Physiol Renal Physiol 2017; 313: F740-55.
12. Akahoshi M, Soda M, Nakashima E, Shimaoka K, Seto S, Yano K. Effects of menopause on trends of serum cholesterol, blood pressure, and body mass index. Circulation 1996; 94: 61-6.
13. Kim JM, Kim TH, Lee HH, Lee SH, Wang T. Postmenopausal hypertension and sodium sensitivity. J Menopausal Med 2014; 20: 1-6.
14. Sudar E, Velebit J, Gluvic Z, et al. Hypothetical mechanism of sodium pump regulation by estradiol under primary hypertension. J Theor Biol 2008; 251: 584-92.
15. Harrison-Bernard LM, Schulman IH, Raij L. Postovariectomy hypertension is linked to increased renal AT1 receptor and salt sensitivity. Hypertension 2003; 42: 1157-63.
16. Dubey RK, Oparil S, Imthurn B, Jackson EK. Sex hormones and hypertension. Cardiovasc Res 2002; 53: 688-708.
17. Scuteri A, Stuehlinger MC, Cooke JP, et al. Nitric oxide inhibition as a mechanism for blood pressure increase during salt loading in normotensive postmenopausal women. J Hypertens 2003; 21: 1339-46.
18. Titze J, Luft FC, Bauer K, et al. Extrarenal Na+ balance, volume, and blood pressure homeostasis in intact and ovariectomized deoxycorticosterone-acetate salt rats. Hypertension 2006; 47: 1101-7.
19. Lara LS, McCormack M, Semprum-Prieto LC, et al. AT1 receptor-mediated augmentation of angiotensinogen, oxidative stress, and inflammation in ANG II-salt hypertension. Am J Physiol Renal Physiol 2012; 302: F85-94.
20. Kong BW, Vanhoutte PM, Man RY, Leung SW. 17β-estradiol potentiates endothelium-dependent nitric oxide- and hyperpolarization-mediated relaxations in blood vessels of male but not female apolipoprotein-E deficient mice. Vascul Pharmacol 2015; 71: 166-73.
21. Reckelhoff JF, Fortepiani LA. Novel mechanisms responsible for postmenopausal hypertension. Hypertension 2004; 43: 918-23.
22. Armando I, Villar VA, Jose PA. Dopamine and renal function and blood pressure regulation. Compr Physiol 2011, 1: 1075-117.
23. Azurmendi PJ, Oddo EM, Obika LF, et al. Gonadectomy influences blood pressure through the kallikrein-kinin system. Kidney Blood Press Res 2009; 32: 342-8.
24. Di Ciano LA, Azurmendi PJ, Toledo JE, et al. Ovariectomy causes overexpression of renal Na(+),K(+)-ATPase and sodium-sensitive hypertension in adult Wistar rats. Clin Exp Hypertens 2013; 35: 475-83.
25. Di Ciano LA, Azurmendi PJ, Colombero C, et al. Defective renal dopamine function and sodium-sensitive hypertension in adult ovariectomized Wistar rats: role of the cytochrome P-450 pathway. Am J Physiol Renal Physiol 2015; 308: F1358-68.
26. Guzik TJ, Hoch NE, Brown KA, et al. Role of the T cell in the genesis of angiotensin II induced hypertension and vascular dysfunction. J Exp Med 2007; 204: 2449-60.
27. Pai AV, Maddox T, Sandberg K. T Cells and hypertension: solved and unsolved mysteries regarding the female rat. Physiology (Bethesda) 2018; 33: 254-60.
28. Chen RY, Fan Y M, Zhang Q, Liu S, et al. Estradiol inhibits Th17 cell differentiation through inhibition of RORγT transcription by recruiting the ERα/REA complex to estrogen response elements of the RORγT promoter. J Immunol 2015; 194: 4019-28.
29. Polanczyk MJ, Hopke C, Vandenbark AA, Offner H. Estrogen‐ mediated immunomodulation involves reduced activation of effector T cells, potentiation of Treg cells, and enhanced expression of the PD‐1 costimulatory pathway. J Neurosci Res 2006; 84: 370-8.
30. Vlachovsky SG, Di Ciano LA, Oddo EM, et al. Expression of proteins related to sodium balance in renal medulla and peripheral blood lymphocytes in a rat model of saltsensitive hypertension. Medicina (B Aires) 2017; 77 (supl 1): 130 (Abstract)
31. Bonventre JV. Dedifferentiation and proliferation of surviving epithelial cells in acute renal failure. J Am Soc Nephrol 2003; 1: S55-61.
32. Kusaba T, Lalli M, Kramann R, Kobayashi A, Humphreys BD. Differentiated kidney epithelial cells repair injured proximal tubule. Proc Natl Acad Sci USA 2014; 111: 1527-32.
33. Sánchez DS, Fischer Sigel LK, Azurmendi PJ, et al. Estradiol stimulates cell proliferation via classic estrogen receptor-alpha and G protein-coupled estrogen receptor-1 in human renal tubular epithelial cell primary cultures. Biochem Biophys Res Commun 2019; 512: 170-5.
34. Márquez LB, Araoz A, Repetto HA, Ibarra FR, Silberstein C. Effects of shiga toxin 2 on cellular regeneration mechanisms in primary and three-dimensional cultures of human renal tubular epithelial cells. Microb Pathog 2016; 99: 87-94.